
 Volume
87, Number 1, January 2006
Volume
87, Number 1, January 2006
Volume
87, Number 1, January 2006
Cover Photo: A Galápagos marine iguana, Amblyrhynchus cristatus, basks in the sun after foraging. Iguanas forage on seaweeds in the intertidal zone during low tide. Foraging ability is reduced by heavy swells and high seas, but even more by the warming effects of El Niño, which reduce nutrient availability and algal growth. Marine iguanas suffered ~50% mortality during the 1982–1983 El Niño episode, and there were substantial reductions during the 1997–1998 event, which occurred while the study was in progress. This photograph was taken in connection with an article by L. R. Vinueza, G. M. Branch, M. L. Branch, and R. H. Bustamente, “Top-down herbivory and bottom-up El Niño effects on Galápagos rocky-shore communities,” to be published in Ecological Monographs 76(1), February 2006.
Table of Contents
(click on a title to view that section)
ANNOUNCEMENTS
Society Notices
ESA 2006 Election Results
Student Awards for Excellence in Ecology
Other Notices
Simon Levin Wins Kyoto Prize
Harvard Environmental Fellowship Award
PHOTO GALLERY: Images from upcoming articles in our scientific journals
Red-eyed Treefrog Metamorphs. K. M. Warkentin
Patch Density Using GIS. H. J. Yoo
Radiotelemetry Used to Study Northern Goshawks. J. D. Wiens
Spadefoot Toads. D. Pfennig
Australian Rangelands. R. McAllister
Ant Parasitism. C. A. Johnson
Post-fledging Survival of Lark Buntings. A. A. Yackel Adams
Fuel Breaks in California Parks. K. Merriam
Galápagos Rocky-shore Communities. L. Vinueza
CONTRIBUTIONS
Commentary
A Review of Theoretical Approaches for Studying the Effects of Interactions between Mutualists and Nonmutualists on Community Stability. M. R. Golinski
Journal Impact Factors. A. A. Agrawal
A Note on “Rising Above the Gathering Storm” (AAAS). E. A. Johnson
A History of the Ecological Sciences, Part 19. Leeuwenhoek. F. N. Egerton
DEPARTMENTS
Focus on Field Stations
The Adirondack Ecological Center at Huntington Wildlife Forest, SUNY College of
Environmental Science and Forestry. S. McNulty
Public Affairs Perspective
Rapid Response Team Update L. Lipps
Society Section and Chapter News
Applied Ecology Section Newsletter
Southeastern Chapter Newsletter
MEETINGS
Meeting Calendar
2006 Midwest Ecology and Evolution Conference
The BULLETIN OF THE ECOLOGICAL
SOCIETY OF AMERICA (ISSN 0012-9623)
is published quarterly by the
Ecological Society of America, 1707 H Street, NW, Suite 400, Washington, DC
20006.
It is available online only, free of charge, at ‹http://www.esapubs.org/bulletin/current/current.htm›.
Issues published prior to January 2004 are available through
‹http://www.esapubs.org/esapubs/journals/bulletin_main.htm›
Bulletin
of the Ecological Society of America, 1707 H Street, NW, Washington DC 20006
Phone (403) 220-7635, Fax (403) 289-9311,
E-mail: [email protected]
|
Associate
Editor Section
Editor, Ecology 101 |
Section
Editors, Section
Editors, |
The
Ecological Society of America
GOVERNING BOARD FOR 2005–2006
President: Nancy B. Grimm, School of Life Sciences, Arizona State University, Tempe, AZ 85287-4501
President-Elect: Alan Covich, Institute of Ecology, University of Georgia, Athens, GA 30602
Past-President: Jerry M. Melillo, Marine Biological Laboratory, Woods Hole, MA 02543
Vice President for Science: Gus R. Shaver, The Ecosystems Center, Marine Biological Laboratory, Woods Hole, MA 02543
Vice President for Finance: Bill Parton, Natural Resource Ecology Laboratory, Colorado State University, Ft. Collins, CO 80523-1499
Vice President for Public Affairs: Rich Pouyat, 3315 Hudson St., Baltimore, MD 21224
Vice President for Education and Human Resources: Carol A. Brewer, Division of Biological Sciences, University of Montana, Missoula, MT 59812-0001
Secretary: David W. Inouye, Department of Biology, University of Maryland, College Park, MD 20742-4415
Member-at-Large: P. Dee Boersma, Department of Zoology, University of Washington, Seattle, WA 98195-1800
Member-at-Large: Shahid Naeem, Department of EEEB, Columbia University, New York, NY 10027
Member-at-Large: Dennis Ojima, Natural Resource Ecology Laboratory, Colorado State University, Ft. Collins, CO 80523-1499
AIMS
The Ecological Society of
America was founded in 1915 for the purpose of unifying the sciences of
ecology, stimulating research in all aspects of the discipline, encouraging
communication among ecologists, and promoting the responsible application
of ecological data and principles to the solution of environmental problems.
Ecology is the scientific discipline that is concerned with the relationships
between organisms and their past, present, and future environments. These
relationships include physiological responses of individuals, structure
and dynamics of populations, interactions among species, organization
of biological communities, and processing of energy and matter in ecosystems.
| Regular member: | Income level | Dues |
| <$40,000 | $50.00 | |
| $40,000—60,000 | $75.00 | |
| >$60,000 | $95.00 | |
|
Student member:
|
$25.00 | |
| Emeritus member: | Free | |
|
Life
member:
|
Contact Member and Subscriber Services (see below) |
Ecological
Applications $50.00 $40.00
Frontiers in Ecology Free to members
Ecological Archives Free
ESA 2006 Election ResultsThe following members have been elected to serve as Society officers, Governing Board members, and committee members. Their terms begin at the end of the 2006 Annual Meeting in Memphis. President (President-Elect August 2006–August 2007, President August 2007–August 2008, Past-President August 2008–August 2009) Vice President for Education and Human Resources (August 2006–August 2009) Members-at-Large (August 2006–August 2008): Board of Professional Certification (January 2006–December 2008) |
REQUEST FOR STUDENT AWARD JUDGES Murray F. Buell Award Judges are needed to evaluate candidates for the Murray F. Buell Award for the outstanding oral presentation by a student and the E. Lucy Braun Award for the outstanding poster presentation by a student at the Annual ESA Meeting at Memphis, Tennessee in 2006. We need to provide each candidate with at least four judges competent in the specific subject of the presentation. Each judge is asked to evaluate 3–5 papers and/or posters. Current graduate students are not eligible to judge. This is a great way to become involved in an important ESA activity. We desperately need your help! Please complete and send this form by mail, fax, or e-mail to the Chair of the Student Awards Subcommittee: Christopher F. Sacchi, Department of Biology, Kutztown University, Kutztown, PA 19530 USA. Call (610) 683-4314; FAX: (610) 683-4854 or e-mail: [email protected] If you have judged in the past several years, this information is on file. If you do not have to update your information, simply send me an e-mail message, “Yes, I can judge this year.” Name ______________________________________________________________________________________________ Areas of expertise (check all that apply): Provide a few key words or phrases that describe your interests and expertise: _________________________
|
|
Murray F. Buell Award and E. Lucy Braun Award Murray F. Buell had a long and distinguished record of service and accomplishment in the Ecological Society of America. Among other things, he ascribed great importance to the participation of students in meetings and to excellence in the presentation of papers. To honor his selfless dedication to the younger generation of ecologists, the Murray F. Buell Award for Excellence in Ecology is given to a student for the outstanding oral paper presented at the ESA Annual Meeting. E. Lucy Braun, an eminent plant ecologist and one of the charter members of the Society, studied and mapped the deciduous forest regions of eastern North America and described them in her classic book, The Deciduous Forests of Eastern North America. To honor her, the E. Lucy Braun Award for Excellence in Ecology is given to a student for the outstanding poster presentation at the ESA Annual Meeting. A candidate for these awards must be an undergraduate, a graduate student, or a recent doctorate not more than 9 months past graduation at the time of the meeting. The paper or poster must be presented as part of the program sponsored by the Ecological Society of America, but the student need not be an ESA member. To be eligible for these awards the student must be the sole or senior author of the oral paper (Note: symposium talks are ineligible) or poster. Papers and posters will be judged on the significance of ideas, creativity, quality of methodology, validity of conclusions drawn from results, and clarity of presentation. While all students are encouraged to participate, winning papers and posters typically describe fully completed projects. The students selected for these awards will be announced in the ESA Bulletin following the Annual Meeting. A certificate and a check for $500 will be presented to each recipient at the next ESA Annual Meeting. If you wish to be considered for either of these awards at the 2006 Annual Meeting, you must send the following to the Chair of the Student Awards Subcommittee: (1) the application form below, (2) a copy of your abstract, and (3) a 250-word or less description of why/how the research presented will advance the field of ecology. Because of the large number of applications for the Buell and Braun awards in recent years, applicants may be pre-screened prior to the meeting, based on the quality of the abstract and this description of the significance of their research. The application form, abstract, and research justification must be sent by mail, fax, or e-mail (e-mail is preferred; send e-mail to [email protected]) to the Chair of the Student Awards Subcommittee: Dr. Christopher F. Sacchi, Department of Biology, Kutztown University of PA, Kutztown, PA 19530 USA. If you have questions, write, call (610) 683-4314, fax (610) 683-4854, or e-mail: [email protected]. You will be provided with suggestions for enhancing a paper or poster. The deadline for submission of form and abstract is 1 March 2006; applications sent after 1 March 2006 will not be considered. This submission is in addition to the regular abstract submission. Buell/Braun participants who fail to notify the B/B Chair by 1 May of withdrawal from the meeting will be ineligible, barring exceptional circumstances, for consideration in the future. Electronic versions of the Application Form are available on the ESA web site, or you can send an e-mail to [email protected] and request that an electronic version be sent to you as an attachment.
Current Mailing Address _____________________________________________________________________________ Current Telephone _________________________________________________________________________________ E-mail __________________________________________________________________________________________ College/University Affiliation ___________________________________________________________________________ Title of Presentation _________________________________________________________________________________ Presentation: Paper (Buell Award) ______ Poster (Braun Award) _______ At the time of presentation I will be (check one): I will be the sole ____ /senior ____ author (check one) of the paper/poster. Signed (electronic signatures are OK) Please attach a copy of your abstract and 250-word or less description of why/how the research presented will advance the field of ecology. |
Other Notices
|
| Inamori Foundation Presents 21st Annual Kyoto Prizes for Lifetime Achievements in Technology, Science, and the Arts American LCD inventor, American ecologist and Austrian conductor KYOTO, JAPAN — November 10, 2005 — Amid the grandeur of the Kyoto International Conference Hall and Japan’s Imperial family, each laureate received a diploma, a Kyoto Prize Medal of 20-karat gold, and a cash gift of 50 million yen (approximately US$425,000) during the prize ceremony, with workshops and lectures continuing through November 12. In addition, the laureates will convene in San Diego, Calif., April 18-20, 2006, for the fifth annual Kyoto Laureate Symposium at San Diego State University; the University of California, San Diego; and the University of San Diego. Basic Sciences Professor Levin’s use of mathematical models to understand the complex patterns of the biosphere has made a substantial impact on environmental sciences and led to new methods of environmental protection. In 1974, with Dr. Robert T. Paine, he proposed “patch dynamics,” the basis of many current ecological models for marine and terrestrial ecosystems. He also demonstrated that high species diversity among competitors -- as observed, for example, in rocky inter-tidal communities, or in tropical rain forests -- can be maintained by recurrent disturbance. Professor Levin has actively collaborated with economists and environmental scientists to propose methods for dealing with environmental problems. His work has shown that ecosystems and the biosphere are not super-organisms, as previously suggested, but complex adaptive systems with apparent regularity emerging from self-organization processes. Among his primary concerns are the staggering losses in biodiversity worldwide that have resulted in the recent past from the mass production, consumption and waste disposal practices of human populations. His 1999 book, Fragile Dominion, illustrates how the loss of biodiversity has created direct threats to human survival, and identifies a series of actions urgently necessary for maintaining biodiversity. In proposing many methods of biological conservation and ecosystem management, Professor Levin has made fundamental contributions to environmental science. Download informative video |
Simon Levin About the Inamori Foundation It is emblematic of the Kyoto Prize that it is presented to individuals or groups in appreciation not only of outstanding achievement, but also excellent personal characteristics. The laureates are selected through a strict and impartial process involving candidates recommended from around the world. As of November 10, 2005, the Kyoto Prize has been awarded to 69 laureates from 12 nations – ranging from scientists, engineers and researchers to philosophers, painters, architects, sculptors, musicians and film directors. The United States has produced the most recipients, with 31 laureates, followed by the United Kingdom (nine), Japan (eight) and France (seven). Kyoto Laureate Symposium In keeping with the philosophy of the Kyoto Prize, the Kyoto Symposium Organization and The San Diego Foundation are sponsoring the 2006-07 Kyoto Scholarships – a program that will assist six high school students (three from the U.S. and three from Mexico) in pursuing college educations within the broad Kyoto Prize fields of Advanced Technology, Basic Sciences, and Arts and Philosophy. Download photos at: http://www.kyotoprize.org |
Harvard Environmental Fellows Program The Harvard University Center for the Environment has created the Environmental Fellows program to enable recent doctorate recipients to use and expand Harvard’s extraordinary resources to tackle complex environmental problems. The Environmental Fellows will work for 2 years with Harvard faculty members in any school or department to create new knowledge, while also strengthening connections across the university’s academic disciplines. The fellowship will provide an annual salary of $50,000 plus health insurance, other benefits, and a $5,000 allowance for travel and professional expenses. Applications and all letters of reference must be received by the Center for the Environment by 15 January 2006. To find out more, visit: ‹http://www.environment.harvard.edu/navigation2/funding.htm› |
|
 |
 |
(Top right) A red-eyed treefrog tadpole and giant water bug, Belostoma sp. This species of treefrog is widespread and locally common in low-elevation forests from Yucatan through Panama. Eggs are attached to vegetation over ponds and swamps, where they are vunerable to arboreal and aerial predators. Hatched larvae drop into the water, where they face aquatic predators. Early hatching is an effective strategy for escaping from several species of egg-eating snakes, wasps, and pathogenic fungi. After hatching, the presence of water bugs overrides the response to fishing spiders, a postmetamorphic predator. (Bottom left) Fishing spiders, Thamasia sp., like this one caught in the act at Ocelot Pond in Panama, prey on metamorphs. Metamorphs emerge later and larger in the presence of these spiders. However, current risk trumps future risk, and tadpoles raised with water bugs do not respond to spiders. |
These photographs were taken in connection with the article by James R. Vonesh and Karen M. Warkentin, “Opposite shifts in size at metamorphosis in response to larval and metamorph predators,” to be published in Ecology 87(3), March 2006. |
Patch Density Using GIS(all rights reserved, used by permission)Click on photos for a larger image. |
 |
 |
 |
| A series of three images demonstrating use of GIS to extract patch density data from large-scale aerial photos of dune vegetation on the Central California coast. Silver dune lupine is the preferred host of western tussock moth at Montaña de Oro State Park, California. Field surveys of the moth matched against spatial data of lupine distribution showed that this poorly mobile moth is more abundant on isolated host plant patches, due to reduced pupal parasitism rates.(Left) A georeferenced aerial photo of a 100 × 150 m area of coastal dune habitat. (Center) A color-classified representation of the photograph, with lupine patches denoted by light-green polygons. (Right) The lupine polygons, filtered and smoothed for noise reduction, and overlaid on the photograph. |
These photographs were taken in connection with the article by H. J. Yoo, “Local population size in a flightless insect: importance of patch-structure-dependent mortality,” |
Radiotelemetry and Northern Goshawks(all rights reserved, used by permission) |
 |
 |
| A radio-marked fledgling Northern Goshawk perched near its nest on the Kaibab Plateau of northern Arizona. Radiotelemetry was used to investigate sources of within- and among-year variation in a study of Northern Goshawk survival. Photo credit: C. Van Cleve.
|
An adult Northern Goshawk closely watches its young on the Kaibab Plateau of northern Arizona. The survival of young Northern Goshawks may largely depend on adult foraging efficiency and defensive behavior. Photo credit: C. Van Cleve. |
Look for the article “Post-fledging survival of Northern Goshawks: the effects of prey abundance, weather, and dispersal,” by J. David Wiens, Barry R. Noon, and Richard T. Reynolds, to be published in Ecological Applications 16(1) February 2006. |
Spadefoot Toads(all rights reserved, used by permission) |
 |
| (Top pair) Plains spadefoot toads (Spea bombifrons) and Mexican spadefoot toads (S. multiplicata) potentially co-occur in the southwestern United States, where their tadpoles develop into either an omnivore morph that eats detritus (left photo) or a carnivore morph that eats fairy shrimp (right photo). In allopatry, both species produce carnivores and omnivores. In sympatry, however, selection to minimize competition has caused S. bombifrons to produce all carnivores, and S. multiplicata to produce all omnivores.
(Lower two pairs) Ecological character displacement such as that between omnivores and carnivores is only possible in ponds where both resources are abundant (upper left photo). In ponds in which detritus is rare (upper right photo), S. multiplicata is absent, and in ponds in which shrimp are rare (lower photos), S. bombifrons is absent. Thus, ecological character displacement, and therefore coexistence of close competitors, is only possible when diverse resources are available. |
These photographs were taken in connection with an article by David W. Pfennig, Amber M. Rice, and Ryan A. Martin, “Ecological opportunity and phenotypic plasticity interact to promote character displacement and species coexistence,” to be published in Ecology 87(3), March 2006. |
Australian Rangelands(all rights reserved, used by permission) |
 |
The sun sets along the road westward to Winton, in Australia’s northern rangelands. Like most rangelands, those in Australia are characterized by a high degree of resource variation, in both time and space. Before Europeans colonized these landscapes, seminomadic humans buffered resource variation through complex social institutions. Pastoralism and private-property rights have since fragmented and disconnected Australian landscapes. But modern Australian pastoral systems are developing their own informal institutions, which are restoring rangeland connectivity over massive spatial scales. |
Look for the article “Pastoralists’ responses to variation of rangeland resources in time and space,” by Ryan J. McAllister, Iain J. Gordon, Marco A. Janssen, and Nick Abel, to be published in Ecological Applications 16(2), April 2006, presenting theory about the Australian perspective on such phenomena. |
Ant Parasitism(all rights reserved, used by permission) |
 |
 |
| A host colony of Temnothorax curvispinosus parasitized by the slave-maker ant Protomognathus americanus and the inquiline ant Temnothorax minutissimus. Both are specialized social parasites, but slave-makers steal the young from host colonies to augment their slave supply. Inquiline ants, having lost the worker caste, spend their entire life cycle within a host colony. The degree of specialization shapes the trajectory of parasite–host coevolution and contributes to the geographic mosaic.
The study by Johnson and Herbers shows that slave-makers P. americanus and Temnothorax duloticus alone negatively affect host colonies, although T. duloticus has a more severe impact. When access to host colonies is shared, the impact is attenuated significantly, suggesting direct asymmetrical antagonism between parasites. In some instances, host colonies host both slave-makers or the slave-maker P. americanus and the inquiline T. minutissimus, indicating a complex multipartite coevolutionary arms race. |
|
These photographs were taken in connection with the article by Christine A. Johnson and Joan M. Herbers, “Impact of parasite sympatry on the geographic mosaic of coevolution,” to be published in Ecology 87(3), March 2006. |
Post-fledging SurvivalPhotographs by A. A Yackel Adams |
 A radio-marked Lark Bunting (Calamospiza melanocorys) fledgling is ready for release on the Pawnee National Grassland, Colorado. Survival estimates that account for age, condition of young, methodological and ecological conditions, and other factors are important for parameterization of realistic population models. |
|
 A radio-marked Lark Bunting (Calamospiza melanocorys) just delivered food to her fledglings, allowing us to estimate survival of young. |
 A female Lark Bunting (Calamospiza melanocorys) at her nest. Adults were easily trapped at nests for radio-transmitter attachment, allowing us to track fledglings. |
 Three-day-old Lark Bunting nestlings begging for food. Young fledge the nest 7–8 days after hatching. Randomly selected young were equipped with transmitters after leaving the nest. Variation in first-year survival can have dramatic effects on population dynamics of passerines, yet is the least-studied component of avian demographics. |
Look for the article by Amy A. Yackel Adams, Susan K. Skagen, and Julie A. Savidge, “Modeling post-fledging survival of Lark Buntings in response to ecological and biological factors,” which will appear in Ecology 87(1):178–188, January 2006. |
Fuel Breaks in California Parks
|

Casper’s Wilderness Park, Orange County, California Federal and state agencies are currently implementing large-scale fuel management programs, including the construction of the fuel breaks shown here in Casper’s Wilderness Park, Orange County, and Santa Monica Mountains National Recreation Area, Los Angeles County, California. |
Fuel break construction and maintenance methods can disturb soils and remove native plant cover, making fuel breaks susceptible to invasion by nonnative plants. Many fuel breaks have significantly higher cover, richness, and density of nonnative species than adjacent wildlands, particularly in areas that have been subject to recurrent fires or grazing. |
These photos were taken in connection with the upcoming article, “Fuel breaks affect nonnative species abundance in Californian plant communities,” by Kyle E. Merriam, Jon E. Keeley, and Jan L. Beyers, to be published in Ecological Applications 16(2), April 2006. |
Galápagos Rocky-shore CommunitiesPhotographs by Luis R. Vinueza |
|
The Galápagos rocky shores provide nesting grounds, food, and shelter to a unique array of organisms, including the only seagoing lizard, the Galápagos marine iguana, Flightless Cormorants, penguins, turtles, and a diverse assemblage of fish, seabirds, crabs, whelks, starfish, and chitons, many of them endemic to the Archipelago.
|
|
 The green sea turtle, Chelonia mydas, screening for food at the interface of the intertidal and subtidal zone. |
 A pair of Galápagos Penguins, Spheniscus mendiculus, resting in the intertidal zone after a foraging trip. Bartolome Island. |
 A pair of Flightless Cormorants, Nannopterum harris, nesting in the intertidal zone of Cabo Douglas, Fernandina Island. |
 A juvenile Yellow-crowned Night Heron, Nyctanassa violacea, looking for crabs in the intertidal, Genovesa Island. |
 A school of yellow-tailed surgeonfish, Prionurus laticlavius, looking for food in the intertidal zone during high tide. |
 The Sally Lightfoot crab, Grapsus grapsus, a very abundant omnivore that feeds mainly on algae. |
 The predaceous crab Ozius verreauxii. |
 The abundant predaceous whelk, Tahis (Nucella) melones, and its prey, the endemic barnacle Tetraclita milleporosa. |
 Marine iguanas and Sally Lightfoot crabs feeding on Ulva in the low intertidal of Genovesa Island. |
Look for the upcoming article by L. R. Vinueza, G. M. Branch, M. L. Branch, and R. H. Bustamente, “Top-down herbivory and bottom-up El Niño effects on Galápagos rocky-shore communities,” to be published in Ecological Monographs 76(1), February 2006. |
A Review Of Theoretical Approaches for Studying the Effects of Interactions Between Mutualists and Nonmutualists on Community Stability |
Introduction The specific goals of this review are threefold. First, I present a brief history of the development of theory for exploring the population dynamics of interspecific mutualism. I then present theoretical approaches that are being used to address the effects of interactions between mutualists and exploiters on community stability. For each approach, I detail results and highlight models that have the potential to be operational in multiple empirical settings. This review can be thought of as preemptive: although the theory is in its infancy, there is a clear distinction between operational and nonoperational models. Without being too bold, I hope this paper leads theoreticians and empiricists toward models that are robust in their predictions, given only a general biological framework within which to work, as opposed to models whose predictions are only relevant for a specific set of a priori assumptions about the biological details of a specific system. 1. Theory development: the effects of intrinsic constraints on the stability of pairwise mutualistic interactions where Ni is the number of individuals, and Ri is the intrinsic rate of growth of species i. The parameter aij describes the negative density-dependent effect of species j on species i. The goal of such models is to track changes in species’ abundances over time. Predictions generated from these types of models are unrealistic, however, because they ignore nonlinearities in per capita growth rates, which when incorporated into the model, give rise to complicated system dynamics far from equilibrium (e.g., unbounded population growth). May (1973, 1974) demonstrated that changes in the population dynamics of pairwise mutualistic interactions could be tracked by changing the sign in Eq. 1 from negative to positive. However, because this model relied on linear functional responses, positive feedback resulted in unbounded population growth of all populations, unless interaction coefficients were small, so that mutualism had little influence on interacting species. Numerous theoretical works have demonstrated that the interaction between two mutualist species can be stabilized by incorporating intrinsic constraints into Eq. 1. For example, the introduction of resource-handling times or metabolic costs incurred by one or both mutualists helps to stabilize the interaction (Vandermeer and Boucher 1978, May 1981, Addicott 1986). Recent mathematical models of population change in pairwise mutualistic interactions have shown that the benefits of mutualism that leads to positive feedback (i.e., saturation of benefits to per capita demographic rates of survival and/or reproduction) do not last forever. For example, by incorporating asymptotically or unimodal saturating functional responses into Model 1, Holland et al. (2002) were able to explain how mutualism has positive feedback, but not indefinitely. Specifically, their model showed that fundamental differences in population dynamics can occur when net effects to that population change linearly, unimodally, or in a saturating fashion. 2. Class 1 models Interestingly, mathematical models for exploring the population dynamics of mutualism in the presence of a third, nonmutualistic species are sparse, and have only recently started to grow in number. This can be attributed to two factors: (1) theoretical ecologists have only recently developed theory for understanding intrinsic mechanisms that stabilize pairwise mutualistic interactions, and (2) the introduction of a third nonmutualistic species into the basic Lotka-Volterra model for mutualism makes interpretation of qualitative dynamics and determination of stability criteria difficult . Although theory has been outpaced by empirical observation, models have been developed to explain the ecology of mutualism in a multispecies community context. Several studies have demonstrated that the incorporation of extrinsic constraints like a third species, either a predator or competitor, acts to dampen mutualist growth (Heithaus et al. 1980, Freedman and Rai 1987, 1988, Freedman et al. 1987). For example, Ringel et al. (1996) built a four-species community model by generalizing Eq. 1 to explore the effects of two additional species, a predator and prey, on the per capita growth of two mutualist species. The effects of each interaction type on the per capita growth of each population was either negative (self and predator effects), neutral (two species that did not interact), positive (prey and mutualism effects), or indeterminate (positive effects for mutualism, negative effects for nectar theft). Results of the model demonstrated that community interactions stabilized the four-species community. However, this type of model did not include intrinsic constraints on the inherent positive feedback between mutualists, which we know to be true in nature. With this in mind, the model of Bacher and Fiedli (2002) is more appropriate for analyzing the dynamics of community stability because their model takes into account interspecific feedback between mutualists. The authors developed deterministic difference equation Lotka-Volterra models to ask the question: How was the well-studied mutualism between the shoot-base boring weevil Apion onopordi and the rust fungus Puccinia punctiformis influenced by the dynamics of their shared host plant Cirsium arvense, and vice versa (Watson and Keogh 1980, Thomas et al. 1994, Fiedli and Bacher 2001). In one version of the model, the rust pathogen was introduced into a thistle population that had reached carrying capacity, and the pathogen was then introduced into a thistle population well below carrying capacity. In the second version of the model, the authors allowed unbounded thistle population growth in order to evaluate the influence of host-plant dynamics on the qualitative dynamics of the three-species system. For both models, intrinsic constraints on the inherent positive feedback between mutualists included a trade-off in benefits of the mutualism for the weevil: ovipositing in infected shoots created an optimal food source for the first generation of offspring, but did not spread the rust, thereby leaving the second generation with an inadequate food supply. Growth of the weevil population increased when feeding on rust-infected plant tissue, which increased byproduct benefits received by the rust, which resulted in infection of healthy thistle in the year following weevil attack. The authors hypothesized that differences in the density of the weevil’s egg load between healthy and rust-infected thistles would regulate population abundances of the mutualists and their host plant. In contrast to the expected hypothesis, both models showed that the population dynamics of the mutualists were largely determined by the dynamics of their host plant. Analysis of the first model showed that neither the starting thistle population size nor its initial carrying capacity had an influence on the size of the thistle population at equilibrium. The second model demonstrated that changes in certain parameter values resulted in unbounded thistle population growth, which resulted in all three species either growing unbounded or going extinct. Hence, three-species coexistence was never reached, which led the authors to conclude that the mutualism between the weevil and the rust alone could not create or maintain system stability. In contrast, the population dynamics of the resource largely determined the stability of the host-plant mutualist. Empirical evidence has demonstrated that exploiters of pollinating seed-parasite mutualisms are present in some communities (Kurdelhue et al. 2000, Pellmyr and Leebens-Mack 2000). Guided by these empirical findings, Morris et al. (2003) developed a mathematical model to explore conditions for stability of an obligate mutualism between a pollinating seed parasite and its plant host, and an exploiter (nonpollinating seed parasite) of the host. The parameters in their model were based on the general biological properties of a well-studied natural system consisting of flowering plants, pollinating seed parasites, and nonpollinating exploiters (Pellmyr and Huth 1994, Kurdelhue and Rasplus 1996, Pellmyr et al. 1996, Pellmyr and Lubens-Mack 2000, Marr et al. 2001). The goals of the model were twofold: (1) One goal was to explore how the rate of interaction between obligate plant-pollinating seed parasite mutualists and nonpollinating exploiters and the type of interspecific and intraspecific competition affected whether the exploiter could invade the plant–pollinator mutualism. (2) The authors also wanted to know whether three-species coexistence was stable. The model was based on the following simplifying assumptions: each fertilized ovule could support at most, one pollinator or one exploiter larva to pupation, and larval survival of exploiter and pollinating seed parasites was reduced by intraspecific competition when more then one egg was oviposited into a single ovule. Thus, the inherent positive feedback between mutualists was constrained by competition. Analysis of the model demonstrated that when competition among and between pollinators and exploiters was weak, the exploiter could invade and the three-species system could persist at a stable equilibrium. However, weak intraspecific competition resulted in a competitive advantage of pollinators over exploiters, which made invasion of the exploiter population and three-way coexistence impossible. 3. Class 2 models Multispecies resource-based population dynamic models Nearly all mutualisms, as is the case with predator–prey and competitive interactions, involve consumer–resource interactions (i.e., the exchange of nutrients between species [Holland et al. 2005]). The chemostat is both a laboratory apparatus and a theoretical construct that is used to investigate the ecological dynamics of consumer–resource interactions in continuous culture. The ecological dynamics of Eq. 1 can be examined within a theoretical chemostat environment, in which the effects of nutrient flow between species can be used to infer consequences of changes in population stability of pairwise predator–prey, competitive, and mutualistic interactions on the stability of communities. In a theoretical chemostat environment, the culture vessel in which the species interact is mixed continuously, which results in a spatially homogenous distribution of nutrients, organisms, and byproducts. Thus, spatial interactions within the vessel can be neglected (Smith and Waltman 1995). Historically, classic continuous-time Lotka-Volterra differential equations have been the mathematical foundation for investigating interactions between predator–prey and competing species in a chemostat. However, the linear functional responses inherent in these equations give rise to unbounded solutions, whereby population sizes of interacting species grow indefinitely (May 1981, Murray 1989, Kot 2001). One solution to this problem is to include asymptotically saturating functional responses in the general Lotka-Volterra model. This allows the largely phenomenological Lotka-Volterra models to include mechanism. Such functional responses are useful because they render unbounded solutions impossible (Smith and Waltman 1995, Kooi et al. 2004) and allow for explicit modeling of nutrient flow through communities (Smith and Waltman 1995, Kooi et al. 2004). Surprisingly, only a limited number of studies have used the chemostat as a theoretical environment for exploring the effects of interactions between mutualists and exploiters on community stability. Kooi et al. (2004) developed a chemostat model of a well-mixed community of two mutualistic prey species with substitutable symbiosis coexisting on a single nutrient, and a predator population that consumed one (specialist) or both (generalist) prey populations. Both prey species were coprophagic mutualists (i.e., each species mutually benefited the other by feeding on the other’s waste products). The authors constrained the inherent positive feedback between mutualists by using a mass-balance model formulation. The model demonstrated that coexistence between a generalist predator feeding on two mutualist populations could only occur if exploitation of the mutualists by the prey was below a critical value. If exploitation was low enough, the predator was able to invade through the boundary equilibrium of two-mutualist coexistence. If exploitation was too great, the community collapsed and all species went extinct. Qualitative behavior of the dynamics of the model were robust to changes in intrinsic species parameters (e.g., intrinsic rate of growth, per capita death rate) and to parameters describing movement of waste products through the system. Spatial–temporal models The current trend in the theoretical study of the effects of mutualist-exploiter interactions on community stability is the use of simulation models incorporating spatial structure. The goal of these types of models is to understand how community stability is driven by the spatial distribution of populations. Empirical studies have demonstrated that there are interspecific mutualisms whose dynamics are regulated by their distribution among habitat patches (e.g., Thompson 1994, Yong et al. 1997, Yu and Davidson 1997, Parker 1999). Empirical studies have also demonstrated that attacks of mutualisms by exploiters in spatially heterogeneous environments can occur in a random pattern over a wide range of spatial scales (e.g., Hawkins and Compton 1992, Bronstein and Hossaert-McKey 1996, Yu et al. 2001). Wilson et al. (2003) used general biological properties derived from these empirical studies to develop an individual-based simulation model that incorporated stochastic interactions and spatial structure. Their model system was composed of two obligate mutualist species and an exploiter species of the mutualism. Like past models, the parameters in their model were based on the general biology of the well-studied interactions between a single species of plant, its pollinator insect, and an exploiter of the plant. However, their model was unique because it incorporated spatial heterogeneity in the distribution of the mutualist and exploiter species. The authors first constructed and analyzed a model of individual-scale interactions between the two obligate mutualist species and an exploiter species. Inherent feedback between mutualists was constrained by adding density-dependent effects in plant reproduction and by adding reduction in seed number as pollinator density became too high. Simulation results demonstrated that incorporation of spatial heterogeneity into the individual-based, nonspatial model resulted in the stable coexistence of all three populations. Analysis of the model demonstrated patterns of temporally constant, fixed-habitat patches. These patterns were robust to changes in the intrinsic parameters of mutualist and exploiter species (e.g., mutualist/exploiter density and life history traits). Bronstein et al. (2003) used a spatial-stochastic simulation model with spatial heterogeneity to explore the population dynamics of obligate, species-specific mutualisms. They based their model on the general biological properties of the well-studied pollinating seed parasite exhibited by fig/fig wasp and yucca/yucca moth interactions (e.g., Dufay and Anstett 2003). Their model system consisted of plants and pollinating seed parasite insects in the presence of one of two obligate exploiter species: florivores or exploiter insects that parasitized seeds but failed to pollinate. Positive feedback inherent to the mutualism was constrained by extrinsic effects of exploitation, rather than the intrinsic effects generated by the mutualist interaction. Simulation results showed that mutualist populations could persist over ecological time in the presence of exploiter specialists, but temporal and spatial dynamics of the mutualist populations were altered in the presence of the exploiter. These patterns were robust to changes in the intrinsic parameters of exploiter species (e.g., exploiter density and life history traits). Conclusions This review has highlighted recent theoretical progress in exploring the effects of mutualist–exploiter interactions on community stability. There are two philosophically distinct classes of models for exploring these effects: (1) bottom-up or individual-case models that are constructed from the biological details of well-studied communities, and (2) top-down or system-based models that are constructed with limited a priori knowledge of the biology of the system. There are many weaknesses in individual case models, among them the fact that they are usually only operational in one particular biological setting, and the fact that their predictions are sensitive to changes in biological details. An additional weakness of the class 1 population dynamic models described in this paper is their use of simplifying assumptions about the biology of communities to make predictions of the model more tractable. This can result in spurious results, because these simplifying assumptions often do not match biological reality. For example, the assumption that only one ovule is produced per plant per season is unrealistic. Most importantly, unlike the predictions of class 1 models, which rely on the inductive method of hypothesis testing, the predications of class 2 models do not rely heavily on a priori knowledge of the biology of a system. Hence, class 2 models are robust in their predictions over a broad range of parameter space, and have the potential to be operational in multiple empirical settings. Acknowledgments Michael R. Golinski is supported by NSF grant DEB-0129630 to Dr. William J. Boecklen. Literature cited Addicott, J. F. 1986. On the population consequences of mutualism. Pages 425–436 in J. Diamond and T. J. Case, editors. Community ecology. Harper and Row, New York, New York, USA. Michael R. Golinski |
CommentaryCorruption of Journal Impact Factors |
Scientists and academic institutions widely use Impact Factors ‹http://wos.mimas.ac.uk/› to evaluate the relative importance of journals. Although sometimes considered controversial, publishing in relatively high Impact Factor journals has been broadly applied as a stamp of approval for hiring and promotions, to rate the accomplishments of academic departments, and the importance of particular disciplines. Both authors and publishers strive to publish high impact journal articles, and the pressure to do so has apparently led to an insidious abuse in how some publishers correspond with authors of nearly accepted manuscripts. At or before the time of acceptance, several journals’ editors are requesting that authors cite additional papers published in that same journal. Some of these requests are general such as ‘We would also appreciate it if you would consider citing relevant past papers [from our journal] in your manuscript’, whereas others are more specific, with journal editors indicating one to several recent (often unpublished) citations. Although the extent of this practice is unknown, at least four major journals in the area of ecology and evolutionary biology routinely encourage such self citation. Because Impact Factors are calculated by dividing the number of citations in the current year (e.g. in 2004) by the total number of articles published in the two previous years (i.e. in 2003 and 2002), citation of articles relatively hot off the press will increase the Impact Factor of a journal. A gentle nudge by an editor to cite additional papers if relevant is all too easy to be uncritically accepted by most authors who are simply overjoyed with the news that their paper has been accepted. To maintain the integrity of objective scientific research, this questionable policy that essentially results in the ‘businessification’ of science must be stopped. Publishers should be embarrassed and authors should not comply. [Reprinted from Trends in Ecology and Evolution, Volume 20, No. 4, Anurag A. Agrawal, Anurag A. Agrawal |
CommentaryThe Gathering Storm |
 In the last couple of years there have been, by my count, no fewer than nine studies in Canada and the United States on science, mathematics, and engineering competitiveness. As is the nature of these reports, they all call attention to the potential for loss of science and engineering innovation, and as a result economic growth and competitiveness. Most of these reports raise three concerns: the increasing importance of non-Western economies, the dwindling number of Western students who wish to go into science and engineering, and the general retreat from teaching the basic sciences in K–12. In the last couple of years there have been, by my count, no fewer than nine studies in Canada and the United States on science, mathematics, and engineering competitiveness. As is the nature of these reports, they all call attention to the potential for loss of science and engineering innovation, and as a result economic growth and competitiveness. Most of these reports raise three concerns: the increasing importance of non-Western economies, the dwindling number of Western students who wish to go into science and engineering, and the general retreat from teaching the basic sciences in K–12.
On 12 October 2005, the National Academy of Sciences of the United States of America released one of the most comprehensive looks at these concerns in a massive document called Rising above the Gathering Storm. The link to the executive summary is ‹http://www.nap.edu/execsumm_pdf/11463.pdf›, and the complete report can be found at ‹http://www.nap.edu/books/0309100399/html/1.html›. The blue ribbon committee that produced The Gathering Storm and the focus groups that assembled the in-depth information provide 10 recommendations. Although The Gathering Storm is a United States document, probably many of the issues and suggestions are applicable in most countries. However, as ecologists, this document offers us another opportunity to remind the public and policy makers that the innovation that will drive the world’s economies in the 21st century will require considerably better understanding of the environment and ecology than has been true in the past. Some of the innovations in past centuries have created the environmental problems of today. E. A. Johnson |
CommentaryA History of the Ecological Sciences, Part 19:
|
There were five outstanding microscopists in the second half of the 1600s: Robert Hooke (1635–1703), Nehemiah Grew (1628–1711), Marcello Malpighi (1625–1694), Jan Swammerdam (1637–1680), and Antoni van Leeuwenhoek (1632–1723). All except Swammerdam had close ties to the Royal Society of London. Leeuwenhoek was the least educated but most persistent of them (Wilson 1995, Fournier 1996, Jardine 1999). The others published their findings during several decades, but by 1700 he was the only one still at it, and he continued this research until his death. Leeuwenhoek’s father was a prosperous basket-maker in Delft who died when Antoni was six. In 1648 Antoni was apprenticed to a cloth merchant in Amsterdam. He returned home about 1654, married, and opened a shop to sell cloth, buttons, thread, and other goods. He became a respected citizen and held several civic posts, and his close contacts included physicians and others better educated than he (Dobell 1932, Schierbeek 1959, Heniger 1973). His next-door neighbor was a physician, Cornelius ‘s Gravesande, who became the city’s anatomist; Leeuwenhoek began attending his dissections in 1668, and in 1681 when Cornelius de Man painted a group portrait entitled “The Anatomical Lesson,” he portrayed Leeuwenhoek standing behind ‘s Gravesande (Leeuwenhoek 1939–1999, III:Plate 1, van Berkel 1982:190–191). |
Fig. 1. Antoni van Leeuwenhoek at middle age. |
Leeuwenhoek saw a copy of Hooke’s Micrographia (1665) and—though he could not read the English text—became intrigued with the illustrations of microscopic investigations. He began making his own lenses and microscopes in 1673, and another Delft physician, Rainier de Graaf, wrote to the Royal Society of London (during the third Dutch–English war) on 28 April 1673 to inform its members that Leeuwenhoek made microscopes that excelled others available. His single-lens microscopes were more powerful than the double-lens ones then in use (Van Zuylen 1982). Along with de Graaf’s note were Leeuwenhoek’s first written observations, which the Society’s first secretary, a German living in London, Henry Oldenburg, translated into English and published in the Society’s Philosophical Transactions.
|
|
|
|
Although Leeuwenhoek published letters in the Philosophical Transactions of the Royal Society of London for the rest of his life, the Philosophical Transactions did not always publish an entire letter, and it omitted publishing a few. Leeuwenhoek wrote some letters on a single topic, but most of them discussed several topics. Collected Dutch editions of his letters began to appear in 1684, and Latin editions in 1685 (Dobell 1932:390–397, Schierbeek 1959:205–209), which included letters that both did and did not appear in the Philosophical Transactions. About a century later, Samuel Hoole translated the published Dutch letters into English (1798–1807), but re-arranged them so that fragments of different letters on the same subject are grouped under topical headings; he did so without dating the fragments. He did provide an index, but no table of contents. Since his translation is reliable (Dobell 1950), I reprinted it in 1977, adding a brief introduction and bibliography. The various editions and translations constitute a bibliographic nightmare, but Francis J. Cole (1937) has provided an excellent guide, both to Leeuwenhoek’s publications and their contents. In 1939 a committee of Dutch scientists began publishing the definitive edition of Leeuwenhoek’s letters—with Dutch and English text on facing pages—and by 1999, 15 of the projected 19 volumes had appeared. Anyone having access to this set of large volumes can dispense with earlier editions for letters written by 1707, but those who lack access to it or wish to consult letters written after 1707 can use Cole’s guide with either Leeuwenhoek’s letters in the Philosophical Transactions or with Hoole’s translation, or both. In a letter to Oldenberg, on 7 September 1674, Leeuwenhoek reported that he had gone to Berkelse Lake and placed some of its water under his microscope. He discovered at least three forms of life: green streaks in a spiral (now called Spirogyra, a green alga), and two kinds of animalcules—apparently what we call rotifers and Euglena viridis. In reporting his discovery of microorganisms, he encountered two problems that he could never handle very precisely: the naming of distinct species and their body parts. Leeuwenhoek was not a draftsman, and his brief verbal accounts aroused both curiosity and skepticism among members of the Royal Society. He hired a draftsman to illustrate his findings, and in 1683 published illustrations of protozoa (Fig. 4), which are of limited interest since the only details are their shapes. |
|
However, by the time he wrote his letter of 9 February 1702, in which he had the (“animalcule”) ciliate Coleps illustrated (Fig. 5), more details were drawn. He had also illustrated a rotifer (Fig. 8), and if not Spirogyra, at least the alga Volvox (Fig. 6). On 14 June 1680, he reported his discovery of the incredibly small animalcules that we call bacteria; later he also had them drawn (Fig. 7). Since the bacteria came from a healthy person’s mouth, he did not think they caused disease. Some biologists have doubted that he could have seen bacteria; however, some of his microscopes still exist and have been used to show that he could have (Ford 1991).
|
|
Leeuwenhoek was an important experimenter (Meyer 1937)—a worthy successor of Redi—though by modern standards his experiments seem quite simple. For example, he discovered minute “vinegar eels” (Turbatrix aceti) in vinegar, and also several other kinds of microorganisms in pepper water (peppercorns submerged in water). Later he added one part of vinegar with “eels” to 10 parts of pepper water and found that all the animalcules in the pepper water died, but the vinegar eels continued to flourish (9 October 1676, Leeuwenhoek 1939–1999, II:125–129). As a merchant, Leeuwenhoek learned to note the size and quantity of goods, and he took this concern into his scientific studies. He developed fairly reliable methods of measurement. He compared the lengths of some microorganisms to the diameter of hairs on cheese mites. To calculate the number of microorganisms in a drop of water, he assumed that a drop of water is the size of a pea, and that a millet seed is 1/91 as large as a pea. He then drew into a pipette a quantity of water the size of a millet seed and divided that amount of water into 30 parts along the pipette, and estimated the animalcules in 1/30 of the water. Finally, he made the calculations to obtain the number in a volume of water the size of a pea. In this case, he estimated there were 2,730,000 animalcules (23 March 1677, Leeuwenhoek 1939–1999, II:119–201). Leeuwenhoek never tired of making discoveries with his microscopes, but he also developed theoretical interests. One of the strongest such interests, which he constantly discussed, was the idea of spontaneous generation of life, against which he collected much evidence, beginning also with his letter of 9 October 1676. Dobell (1932:136, note 1) interpreted some comments in this letter as showing Leeuwenhoek debating with himself the possibility of spontaneous generation before he reached his firm opposition to the idea seen in his later letters, and Ruestow (1984) agreed. However, Leeuwenhoek’s modern editors think this is a misunderstanding of Leeuwenhoek’s less than crystal-clear comments (in Leeuwenhoek 1939–1999, II:101), and Smit (1982) appears to agree with these editors, since he does not discuss any ambivalence in this letter. The occasion for Leeuwenhoek raising the question of spontaneous generation was his discovery of microorganisms in rainwater that had stood in a cask for several days. A later example of Leeuwenhoek’s observations discrediting spontaneous generation is in a letter written 9 February 1702, stating that on the previous 25 August he had found animalcules (rotifers) in water from a house gutter, in which he observed that they became dry for a time, and then when wet again, their bodies swelled and they swam off. He thought that if one did not know that they were dormant in the dry matter and then it became wet, one might think they arose spontaneously in the wet matter. Another of Leeuwenhoek’s strong theoretical interests was in reproduction. This interest was sharpened by the discovery of spermatozoa (“animalcules” to him). A medical student, Johan Ham, told him in the summer of 1677 about his discovery of animalcules in the semen of a man with venereal disease; he believed they arose from the putrefaction of the semen. Leeuwenhoek refused to accept Ham’s idea on their origin, since that implied spontaneous generation. His study of his own semen (from his marriage bed, he informed the Royal Society) showed that spermatozoa are natural to semen. He reported his observations to the Royal Society in a letter written in November 1677, and in a subsequent letter of 18 March 1678 he included drawings of both human and dog sperm (Leeuwenhoek 1939–1999, II:280–293, 346–349, Plates 16–17). During his lifetime, he described sperm from 30 kinds of animals: 7 mammals, 2 birds, 1 amphibian, 7 fish, 11 arthropods, and 2 mollusks (Cole 1937:8). He soon concluded that sperm are embryos, and he rejected his townsman de Graaf’s conclusion that embryos arise after intercourse when eggs from mammalian ovaries enter the Fallopian tubes (Lindeboom 1982, Ruestow 1983). Leeuwenhoek thought ovaries are nonfunctioning in females, just as nipples are nonfunctioning in males. In a letter dated 13 June 1679, he rejected an Aristotelian report that mice reproduce by parthenogenesis (Leeuwenhoek 1939–1999, III:73–83), but in a long letter written on 10 July 1695, he reported (Leeuwenhoek 1939–1999, X:269–301) his discovery of parthenogenesis in aphids. He returned to this subject in subsequent letters (Cole 1930:90, 1937:224, Egerton 1967:6–16). After discovering parthenogenesis, he should have rethought his belief that embryos are contained in sperm. On other occasions he admitted his mistakes, but concerning parthenogenesis he went to extremes to avoid doing so (Cole 1937:12–13, Schierbeek 1959:105–106). Still another of Leeuwenhoek’s persistent interests was in parasites. We saw in Parts 17 and 18 (Egerton 2005b, c) that this interest was fairly common among contemporary naturalists and physicians—focused on the natural history of parasites and interactions with hosts, without the parallel development of a theory of parasitology. In those days, most people had some first-hand knowledge of fleas and lice, and we have already seen that in his first letter to the Royal Society he had included observations on louse anatomy. He reported further on lice in a letter dated 15 February 1677, which is lost. In his letter of 5 October 1677, he reported observations on the development and metamorphosis of fleas. He had put several in a container and found that a flea can lay 15 or 16 eggs in 24 hours. He then carried enclosed eggs in his pocket and found they hatched in 8 or 9 days. He described the external anatomy of a larva and compared it to that of silk worms. He thought Swammerdam had mistaken flea droppings for eggs (Leeuwenhoek 1939–1999, II:245–253). Neither of these microscopists distinguished the different species of fleas they studied (Van Bronswijk 1982). In autumn Leeuwenhoek observed larvae spin cocoons, and a few days later he opened some cocoons and found inside weak fleas, which he thought were affected by the cold, indicating that they would not have come out by themselves until the winter ended (14 January 1678, Leeuwenhoek 1939–1999, II:319). On 12 November 1780, he sent observations on flea sperm (Leeuwenhoek 1939–1999, III:327). He reported further on flea anatomy and physiology in letters of 22 January 1683, and 15 and 27 October 1693. A goal of his flea studies was to determine the time period for each stage in the life cycle from egg to adult, which he finally achieved in his letter of 15 October 1693 (Leeuwenhoek 1939–1999, ?:211–227). He allowed fleas to suck blood from his hand in order to see the effect of food on egg laying. |
|
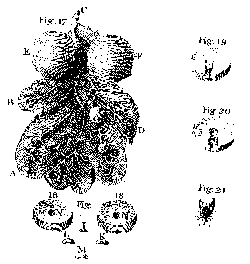 Fig. 9. Oak leaf with galls, cross-sections of a gall, four larvae, and adult fly (Leeuwenhoek, Hoole I:Plate 5, Figs. 17–21). |
|
| Leeuwenhoek examined flatworms (flukes) from the livers of diseased sheep under a microscope and suspected that the sheep got the worms from drinking rainwater that collected in fields (21 February 1679, Leeuwenhoek 1939–1999, II:417–419). He pursued the subject no further until 1698, when he and Professor of Medicine Goderfridus Govard Bidloo (1649–1713) of Leiden University (van der Pas 1978) discussed liver flukes in sheep. Both then wrote up their observations for publication, with Leeuwenhoek sending his to the Royal Society and Bidloo sending his to Leeuwenhoek, who had them published in Delft. Bidloo sent with his letter an overly precise drawing of a fluke, which shows two eyes, a heart, a circulatory system, and intestines that existed only in his imagination. Nevertheless, Bidloo did recognize the eggs and concluded correctly that the species is hermaphroditic. He also generalized from his observations that these worms seem to cause disease in sheep and that worms probably also cause disease in humans (Bidloo 1698, 1972). Leeuwenhoek went out and attempted to find fluke eggs in fields and ditches, where they might have been deposited in sheep feces (2 January 1700, 1939–1999, ?), but he had no way to identify them if he had found them. The fluke life cycle is so complex that it was not fully understood until the mid-1800s (Reinhard 1957). | |
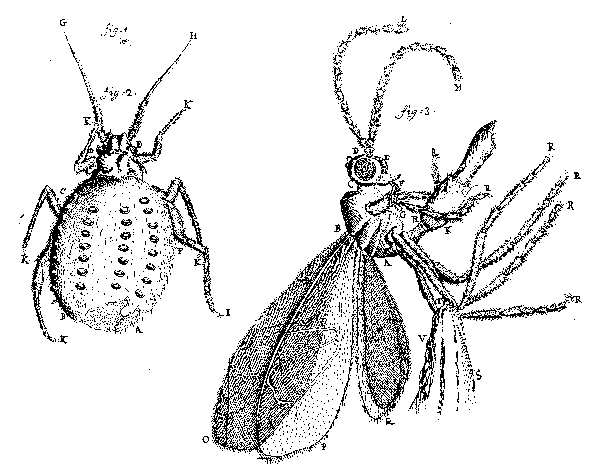 Fig. 10. Leeuwenhoek’s Fig. 1 is a “green louse” (aphid) natural size; his Fig. 2 is an aphid shell seen under a microscope, from which a fly had emerged at the bottom; his Fig. 3 is a parasitic fly that emerged from an aphid (26 October 1700, Royal Society of London Philosophical Transactions 22:facing p. 655). |
|
| Once when Leeuwenhoek had loose stools, he examined his feces under a microscope and described to the Royal Society the microorganisms he found (protozoa and spirochaetes or Spirillum), and he did not find them in his feces when he did not have diarrhea (4 November 1681, Leeuwenhoek 1939–1999, III:367–371), but he drew no conclusion about animalcules causing diarrhea.
In Holland, “gall-nuts” were imported from Aleppo, Syria for making dye. Leeuwenhoek assumed from the name that they were actually nuts, until he saw a local variety on oak trees and realized that they must be stimulated by an insect. These galls were formed upon the large fibers, or vessels in the leaves, which were burst or broken, in the places where the galls were formed; so that I concluded that some insect had wounded or gnawed those vessels, and that the juices of the tree, flowing out of the wounded part, had extended themselves in globules and vessels, and thus, at length caused the formation of the gall-nut. [14 May 1686, Leeuwenhoek 1977, I:137] He cut open some galls and found inside a living white worm. By continuing to open others periodically, he discovered that the worms became flies. He also studied “thistle-nuts,” which people carried in their pockets as health charms. He had a draftsman illustrate both kinds of galls and the associated insects (Fig. 9). In 1700, fruit trees in Delft were infected with a great many flies, but when Leeuwenhoek examined them, he found they were associated with even more green lice (aphids), whose parthenogenesis he had discovered in 1695. The flies laid their eggs in the aphids, and later flies emerged from an aphid shell (Fig. 10). |
|
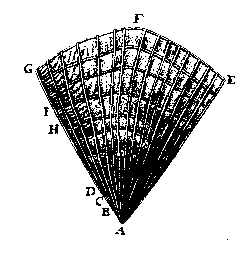 Fig. 11. Leeuwenhoek’s cross-section drawing of an oak trunk was given as a pie-shaped wedge rather than in full circle as is the modern custom; he said the oak tree was 12 years old and 4 2/6 inches in diameter (written 10 July 1686, but not published until the September–October 1694 issue of the Royal Society of London Philosophical Transactions 18:facing p. 193). |
|
| Leeuwenhoek wanted to know not only the size and quantity of organisms he studied, but for some he eventually wanted to determine their age. He first explained briefly at the end of his letter dated 12 January 1680 (Leeuwenhoek 1939–1999, III:185) the use of annual rings to determine the age of trees, and six years later he discussed it again and sent the Royal Society an illustration of a tree seen in cross-section (Fig. 11). We saw in Part 18 (Egerton 2005c) that Ray explained age determination in trees in his Cambridgeshire flora (1660), but Leeuwenhoek would have been unable to read that book in the unlikely event that he ever saw it.
He became interested in fish scales initially because Jews thought they could not eat eels and burbot, because each supposedly lacked scales and was therefore forbidden by Scripture. Using a microscope, he showed that they do have scales. When he examined an individual scale under a microscope he saw concentric dark lines, which he interpreted correctly as annual rings. The scale that he had drawn was thus seven years old (Fig. 12), and he assumed that this was also the eel’s age. We now know, however, that eel scales only appear at age three, and therefore the eel would have been age 10 or 11 (Leeuwenhoek 1939–1999, IV:293–297, and note 48). Later, he attempted to determine the age of other fish, and an elephant’s tooth (Egerton 1967:9). When he turned to shellfish, he discovered that the layers of the shells were too numerous to be annual rings, and he speculated that they were laid down monthly, with the new moon (Palm 1982:159). |
|
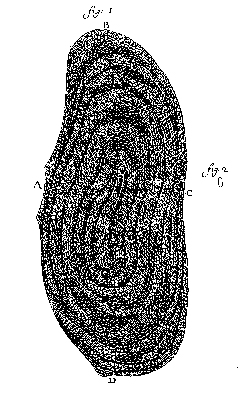 Fig. 12. Eel scale; Leeuwenhoek’s Fig. 1, as drawn with a microscope, and Fig. 2 represented the actual size (written 25 July 1684, Royal Society of London Philosophical Transactions 15: facing p. 883 (1685) or Leeuwenhoek 1939–1999, IV:Plate 27). |
As a businessman and prominent citizen, Leeuwenhoek had an interest in the practical application of his investigations, and many of them were undertaken to clarify practical problems. His studies of insect life histories are examples (Bodenheimer 1928–1929, I:367–379, II:363–367, Schierbeek 1959:Chapter 6). His study of the grain weevil Calandria granaria in 1687 had the dual motive of studying an important pest, and also providing another opportunity to discredit the idea of spontaneous generation. On 13 March, he obtained some calandars and put 6, 8, or 9 in three vials along with 6, 10, or 12 wheat grains and carried them in a leather case in his pocket. He saw them mate on 27 March, and discovered that they lay few eggs. Comparing this to silkworms, which lay many eggs in one or two days and then die, he concluded that calandars must live longer as adults in order to lay eggs several times. He saw that females lay only one egg in a wheat grain, and he suspected that frequent stirring of stored wheat could prevent them from depositing their eggs (Leeuwenhoek 1939–1999, VII:31–33). |
| In the same year he also studied reproduction in a fly, probably Calliphora erythrocephala, which laid about 144 eggs. He followed the progress of eggs laid on 9 September and found that they emerged from pupae as adults on 12 October. He then calculated that the theoretical rate of increase over three months, assuming no mortality, resulted in 746,496 flies (written 17 October 1687, Leeuwenhoek 1939–1999, VII:81–133). This was an important step for animal demography, the first example of what Royal Chapman much later called “biotic potential” (1931:182), and Birch called “the intrinsic rate of natural increase of an insect population” (1948). Later, he calculated the potential rate of increase for other species, and speculated on factors that limit their increase, usually food or climate (Egerton 1967:14–19). He was also one of the earliest investigators of a food chain—an aquatic one that involved haddock eating shrimp and cod eating haddock (10 September 1717, Leeuwenhoek 1798–1807 and 1977, I:283–285). In an earlier letter (2 June 1700, Leeuwenhoek 1939–1999, XIII:92–95) he had discussed what shrimp ate, but he did not link that information to his 1717 letter.
Leeuwenhoek lived almost 91 years and devoted the last 50 of them to science, primarily to biology, with an impressive number of his discoveries being on natural history, many of these on what we call ecological topics. His research and publications made him famous throughout Europe, and he was highly esteemed by leading scientists of the time. Acknowledgments I thank Jean-Marc Drouin, Muséum National d’Histoire Naturelle, Paris, and Anne-Marie Drouin-Hans, Université de Bourgogne. |
|
| Frank N. Egerton Department of History University of Wisconsin-Parkside Kenosha WI 53141 E-mail: [email protected] |
|
The Adirondack Ecological Center at Huntington Wildlife Forest, State University of New York College of Environmental Science and ForestryThe Adirondack Ecological Center (AEC) is a field station located on the Huntington Wildlife Forest (HWF), The Adirondack Mountain region is of national and global significance because it is a relatively intact natural ecosystem encompassing 2.5 million hectares and contains much of the remaining old-growth forest and wilderness in the eastern United States. Creation of the Adirondack Park in 1892 and subsequent state legislation brought about an unusual suite of regulations for public and private land that was designed to protect this ecosystem. However, the potential impacts of human activities on the region are immense. The Adirondack Mountains are threatened by development and pollution: the region is within a day’s drive of nearly one–third of the North American human population, and development is heavy along roadsides and lakeshores. The size of the region, diversity of ecological communities, natural resource-based economies and legal protection make this region an ongoing experiment in sustainability and therefore of vital interest to science and society. In 1932, Archer and Anna Huntington established HWF for what was then the New York State College of Forestry at Syracuse now known as ESF (Fig. 2). HWF is located in the Town of Newcomb, Essex County and the Town of Long Lake, Hamilton County (44°00' N, 74°13' W). HWF was established specifically to provide an experimental site on which to investigate the central Adirondack ecosystem. The Adirondack Ecological Center (AEC) was established at HWF in 1971 with the intent of bringing more scientific expertise to the Adirondack Mountains and a long-range goal of broadening the impact of research and educational programming. ESF has conducted research and educational programming at HWF for nearly 75 years. Data collected through monitoring and research projects are incorporated into the Adirondack Long-Term Ecological Monitoring Program (ALTEMP). ALTEMP provides an institutional framework for monitoring of over 100 physical, chemical and biological variables. Recognizing the value of these long-term datasets, HWF was chosen as a study site for the National Atmospheric Deposition Program (NADP), National Trends Network (NTN), Mercury Deposition Network (MDN), and Clean Air Status and Trends Network (CASTNET). AEC joined the Northeastern Ecosystem Research Cooperative (NERC), an initiative encouraging scientific collaboration in the U.S. and Canada, in 2000. |
|
HWF is located in the transition between boreal and deciduous forest. Northern hardwood stands include American beech (Fagus grandifolia), sugar maple (Acer saccharum), red maple (Acer rubrum), and yellow birch (Betula alleghaniensis). Conifer stands consist of red spruce (Picea rubens), and balsam fir (Abies balsamea) at higher elevations and along water courses. Mixed hardwood-conifer stands contain yellow birch, red maple, red spruce, and eastern hemlock (Tsuga canadensis), often associated with white pine (Pinus strobus) or Northern white cedar (Thuja occidentalis). Soils in the study area are primarily glacial till and elevations range from 457 to 1403 m. Mean monthly temperatures range from –10°C in January to 18°C in July, and annual precipitation averages 100 cm. Nearly 300 vertebrate species are found on HWF, including almost 200 birds, black bear (Ursus americanus), white-tailed deer (Odocoileus virginianus), recently returned moose (Alces alces), and a host of small mammals and herpetiles. Mission The mission of the AEC is to provide an understanding of the Adirondack ecosystem through research. The breadth and depth of research conducted at HWF provides material for educational programming. The AEC supports the broader mission of ESF by providing opportunities for students to gain direct experience with scientific research and natural systems. Student experiential learning bridges the gap between conceptual classroom learning and application of knowledge to problem-solving. |
|
 Figure 3. Arbutus Cabin. |
Facilities
HWF operates year-round; all buildings have electric heat and light and some have kitchen facilities. Housing facilities in two areas on HWF currently accommodate up to 83 people during summer months. The Arbutus Lake Area contains researcher and graduate student housing and is often used for small classes and conferences. Several of these buildings were originally used by the Huntington family as a vacation home or “Adirondack Great Camp” (Fig. 3). They were built using plans of the famous Great Camp designer, William West Durant, and have significant historic character. Huntington Lodge has housing for 17 people, while other cabins in the area house 4-6 people each. |
| The Rich Lake Area contains the Multipurpose Building, Director’s Residence and the White Pine Bunkhouses. The Multipurpose Building provides a cafeteria-style dining center, bathrooms, showers and laundry (Fig. 4). Two bunkhouses hold 32 people (2 rooms with 8 bunks each) and are used for student or short-term housing. Bunkhouses are within walking distance of the Multipurpose Building and include handicapped-accessibility. By 2006 two additional bunkhouses with room for 32 will be completed.
The AEC office building contains office space, a computer lab, conference room, library, and storage facilities. The lab includes computers, color and large-format printers, and high-speed internet access. Computers are networked and a wireless network allows roving laptops to connect throughout the building. The conference room is equipped with LCD projectors and a videoconferencing/distance learning system and is capable of seating 50 people. The AEC library holds approximately 1500 titles, including all graduate theses and dissertations relating to work on HWF, as well as technical and popular journals, textbooks, and other reference materials. The AEC library is integrated into the ESF/Syracuse University libraries, searchable via the Internet. The AEC maintains a Geographic Information System as well as a collection of about 500 museum specimens of birds and mammals for teaching purposes and an herbarium of 1,152 species that serves as a reference collection. The Animal Studies Laboratory provides laboratory space, a wet lab, freezers, drying ovens, and cages for holding captive animals up to 20 kg in size. An outdoor pen complex for holding captive animals is also associated with this building. The Maintenance Garage contains workspace for carpentry and vehicle maintenance including a hydraulic lift. There are a variety of storage buildings throughout the property. HWF maintains snowmobiles, all terrain vehicles, 4-wheel drive trucks, and boats. A variety of field research equipment is available and includes animal traps, vegetation survey instruments, limnological and fisheries gear, and climatological and hydrologic devices. There are Trimble and Garmin GPS units available. The research portion of HWF is gated and has 35 km of gravel roads; 3 km are plowed in winter and the rest are snowmobile-accessible. Approximately 400 ha area of old-growth forest occurs on HWF and was designated by action of the ESF College Trustees in 1941 as the Natural Area. The Natural Area serves as a control area for long-term studies. Of the seven lakes on HWF, Wolf Lake is rated as among the most pristine in the Adirondack region due to an assemblage of native aquatic species and limited influence by invasive species or human activities. |
|
 Figure 4. The Multipurpose Building and Dining Center. |
|
| The weather station has operated continuously since the 1930s. A gauging station was installed in 1989 on Arbutus Lake, a watershed at the headwaters of the Hudson River drainage; today this is the only continuously monitored gauge in the Adirondack Mountains. In 1990, a 38-meter tall walk-up tower with meteorological instrumentation for sampling air chemistry was installed by the Environmental Protection Agency and used by the National Oceanographic and Atmospheric Administration as part of the Atmospheric Integrated Research Monitoring Network (AIRMoN). Meteorological measurements were significantly upgraded in 1991 with the installation of an automated system to monitor temperature, relative humidity, solar radiation, barometric pressure, rainfall, and wind speed and direction. Air and water temperature and stream discharge data are collected with data loggers at the Arbutus Lake Outlet (gauged with a V-notch weir) and communicated in real-time to ESF researchers in Syracuse. The Arbutus Lake Inlet has been monitored since 1994 with an H-flume equipped with an automated discharge logging collection system. In addition, solute chemistry is sampled with piezometers, water table wells, soil tension lysimeters, snow lysimeters and throughfall collectors installed throughout the Arbutus Lake Watershed. The EPA CASTNET installation provides atmospheric data on the dry deposition component of total acidic deposition and ground-level ozone. Seismic activity is measured and telemetered to Columbia University. A wireless communication system that will allow remote transmission of data and real-time access to the hydrological, meteorological, and biochemical research at this site is expected to be online in 2006.
Staff at the AEC include a Director, Associate Director, administrative assistant, research support biologists, educational program coordinator, GIS specialist, forest properties staff, maintenance staff, and Dining Center staff. The Webb and Sage Apprenticeships are opportunities to provide a current undergraduate student or recent graduate with professional experience. The apprentices gain experience by serving as a full member of the staff while developing skills in field biology, research, and leadership. Research assistants at the graduate and undergraduate levels are employed during summer. Key attributes of HWF are its size and security. HWF offers the ability to conduct experimental manipulations on scales of tens to thousands of hectares. Approximately 85% of the 6,000 ha are behind locked gates accessible only by permission. HWF is large enough to accommodate experiments on areas of 10 to 2000 hectares and allow assessment of ecological effects across multiple spatial scales and forest types. In addition, agreements allow investigator access to adjacent state and private land parcels with different land-use histories. Many researchers use HWF as one of a network of sites to study ecosystem processes along a land use, forest age, or forest management gradient. |
|
 Figure 5. Students check a salamander research plot. |
Research
Research topics pursued at HWF are generally focused on ecology, hydrology, biogeochemistry, mammalogy, limnology, silviculture and wildlife management (Fig. 5). Two broad research areas encompass most of the research activity at HWF: Integrated Forest Ecology and Management. Investigations focus on the dynamics of northern hardwood forest, alternatives for forest management, and the interaction of forest vegetation and vertebrate populations. |
| Key facets of this work include development of both even and uneven-aged silvicultural methods, understanding of the role of vertebrates in shaping forest species composition, the importance of understory plants and vertebrates as indicators of biotic integrity, and the recognition of the long-term effects of event-driven processes. Hypotheses developed at HWF are now being tested across the Adirondack Mountains and in other areas of the Northern Forest (northern Maine, New Hampshire, Vermont, and New York).
Biogeochemistry. Biogeochemical studies at HWF are part of regional and international networks and have an emphasis on forest ecosystems and freshwater. Research has included evaluations of atmospheric deposition, and investigations of the role of nitrogen and sulfur in soils and watersheds. These biogeochemistry studies have promoted cooperation among scientists in the U.S. and worldwide. Today, a multidisciplinary approach draws scientists at ESF, Cornell University, Institute of Ecosystem Studies, Kyoto University, Oregon State University, Syracuse University, United States Forest Service, United States Geological Survey, University of Calgary, and University of Waterloo. Studies at HWF are integrated with Hubbard Brook Experimental Forest in New Hampshire, Croton Watershed of New York, Bear Brook Watershed in Maine, Oneida Lake in New York, and Lake Erie research. Among the most important research findings of recent years are the following: Forest Dynamics: A series of studies dating back to the 1950s culminated in a new understanding of the interaction of herbivores, disease pathogens, site conditions and weather in shaping the species composition of northern hardwood forests. This work offers an alternative explanation to classic succession theory and gap-phase dynamics for the origin of the current overstory: the composition of the forest canopy persisting over centuries in this environment is shaped largely by episodic disturbance events occurring within a context of co-incident states of multiple ecological factors. Measuring Biotic Integrity: An issue of debate in the Adirondack Park has been how to measure the influence of human activity on the integrity of natural ecosystems. Research conducted through the AEC adapted the concept of biotic integrity from aquatic science to terrestrial environments. The index is built on breeding bird distributions across the region, and specifically, bird guild representation. Findings show that a relatively simple model incorporating landscape variables such as density of road networks can effectively predict changes in guild structure. Biogeochemistry: Atmospheric deposition is an issue of concern throughout the eastern U.S. Research at HWF over the past 20 years has documented important changes in nitrogen and sulfur levels. A key finding is that movement of ions is more complex than originally hypothesized and highly dependent on not only local soil but also plants. For instance, the presence and role of nitrogen-fixing shrubs in wetland ecosystems are a critical regulating influence on the biogeochemical dynamics of nitrogen. Forty-five research projects were conducted on HWF in 2004–2005. Between 100–125 faculty, agency scientists, graduate students and undergraduate research interns are active each year. Researchers are from ESF and other institutions including the State University of New York Colleges at Buffalo, Cortland and Potsdam; Columbia University; University of California-Davis; Cornell University; USGS; EPA; NOAA; US Bureau of Mines; and US Forest Service. Over the 70-plus year history of HWF, 500 refereed publications and 160 theses and dissertations have been completed. At the outset of an investigation, a Research Study Plan must be filed and approved. Summaries of current research are found at www.esf.edu/aec/research/currentresearch.htm. Data are stored in indexed file cabinets and on electronic media. An on-line publications list (www.esf.edu/aec/publications/publications.htm) is searchable by author and category. When permitted by publishers, abstracts are available on line. |
|
| Education
Results of research at the AEC have fed directly into formal educational programs. Over 200 students took college courses in 2004–05, in courses such as Environmental Interpretation, Aquatic Entomology, Adirondack Forest Ecology and Management, Winter Mammalian Ecology, Forest Health, and Limnology & Fisheries (Fig. 6). Each year, HWF hosts numerous field trips with roughly 275 students attending from various academic institutions. |
 Figure 6. Mammalogy capture and handling class. |
| While college students are the primary focus of educational activities on HWF, several instructional programs focus on 7–12th grade students. Students with diverse cultural backgrounds are brought into contact with natural systems, sometimes for the first time, and introduced to ecology, the scientific process, and college life. Approximately 320 students participate in these programs.
AEC also seeks to broaden the educational impact of Adirondack research through dissemination of scientific information to the general public. In 1990, AEC partnered with the state Adirondack Park Agency to build a Visitor’s Interpretive Center (VIC) on HWF. The VIC houses dioramas, classroom and office space, and an auditorium with a capacity of 150 people. Associated with the VIC are 4 km of trails with interpretive signs that introduce people to upland and lowland forest, wetland and lake communities. Each summer the AEC sponsors the Huntington Lecture Series at the VIC; the series attracts 350 visitors annually. In 1995, AEC partnered with the Town of Newcomb to upgrade an historic fire tower, construct a public parking area and trailhead on Goodnow Mountain, and create a kiosk and a series of interpretive educational booklets that expose visitors to research outcomes. The number of people accessing the VIC and the Goodnow Trailhead is over 25000 per year. Visiting researchers and courses Workshops, conferences, and short courses for professionals attract over 250 enrollees each year. For instance, each year 25 high-school biology teachers are exposed to current research methods and topics through Stalking Science Education in the Adirondack Mountains, an intensive weeklong course. The course focuses on the process of science and shows teachers how to use outdoor environments to excite students about science. An NSF-sponsored Chatauqua Short Course in Mammalian Ecology is offered to college science teachers in undergraduate education. Continuing education workshops in forest and wildlife management are geared toward natural resource professionals who wish to pursue certification. In 2004–05, 7,200 user days were logged at HWF, including research, class and field trip use. With new facilities coming online, there will be more space for research and educational use of HWF. Visit HWF online at http://www.esf.edu/aec or for more information contact: Dr. William F. Porter Stacy McNulty |
|
Public Affairs Perspective |
  |
Applied Ecology Section NewsletterHistory and mission The Applied Ecology Section of the Ecological Society of America is the second largest and the third oldest of the active sections within the Society. The Section was established in 1971 and has a twofold purpose: 1) To facilitate communication about applying ecological principles to solve practical environmental problems, and Call for nominations for Section officers for 2006–2007 Article 5. OFFICERS. The officers of the Section shall be a Chair, Vice-Chair, and Secretary. The Officers shall comprise the Section Executive Committee and may act on behalf of the section during intervals between annual meetings. Voting for Officers shall be by either mail or by email ballot distributed to members in odd numbered years. Officers shall serve for a term of two years and not be eligible for re-election. The Chair and Secretary assume office in the year the election is held, and the Vice-Chair assumes office the following year. Article 6. CHAIR. The Chair shall preside at the business meetings of the Section, authorize expenditures of Section funds, and shall promote in every practical way the interests of the Section. The Chair shall appoint a Nominating Committee, which shall prepare a slate of candidates for each office. Article 7. VICE-CHAIR. The Vice-Chair shall be responsible for arranging the scientific program for all meetings of the Section, and shall assume the duties of the chair whenever that person is unable to act. Article 8. SECRETARY. The Secretary shall keep the records of the Section and an up-to-date membership and mailing list, and shall perform such other duties as may be assigned by the Chair. Duties that may be assigned by the Chair can include: submits information to ESA Bulletin by 15 January, April, July, and October for publication in the next ESA Bulletin; takes minutes at the annual meetings; maintains and updates the web page; assists the Chair and Vice Chair with distributing information and other tasks as deemed by the Chair; assists with organizing and tallying votes for the Student Travel Award; counts ballots for elections. Please send nominations, including a one-paragraph biosketch that describes your vision as an Officer of the Applied Ecology Section, by 15 May 2006 to the Nominations Chair: Becky Kerns, Research Ecologist Officers for 2004-2006 Co-Chairs: Vice Chair Secretary Student Travel Awards The Applied Ecology Section will support students in their efforts to present their work at the 91st ESA Annual Meeting in Memphis, Tennessee, 6-11 August 2006. The Section is now calling for nominations for scholarships, with individual awards up to $750 depending on the funding available and number of qualified applicants. The deadline for receipt of applications is 15 May 2006. Instructions and details for the application process can be found at the Applied Ecology Section web page at ‹http://www.esa.org/applied/› Last year’s scholarship recipients were able to present their research at ESA’s 90th Annual Meeting in Montréal. The amount of each award and the name and address of each student are listed below. Susan Tilley’s award came from the Applied Ecology Section’s long-range planning grant, which was awarded on 21 December 2004. The other awards came from the Section’s overall funds.
Long-Range Planning Grant The Applied Ecology Section received a long-range planning grant from the ESA Governing Board to help implement ESA’s Long Range Plan. The award was for $1000, and was used for student scholarships, webmaster training, and the business mixer at the 2005 Annual Meeting. A written report on how the funds are used was presented to the Governing Board in Montréal, Canada. Annual Meeting update The Applied, Agroecology, Rangelands, and Long-Term Studies Sections held a joint mixer during the 90th ESA Annual Meeting in Montréal, Canada, on Wednesday, 10 August 2005. The Applied Ecology Section held its business meeting on Tuesday, 9 August 2005 during the 90th ESA Annual Meeting. Attendees included: Marty Spetich, co-chair; Becky Kerns, vice-chair; Hal Balback, U.S. Army Engineer Research and Development Center, Champaign, Illinois; Paula Armstrong, consultant, Canada; and Marty Matlock, Associate Professor of Ecological Engineering, Fayetteville, Arkansas. Southeastern Chapter Newsletter
|
2006 Midwest Ecology and Evolution Conference (MEEC)The Conference will be held 17–19 March 2006, hosted by St. Louis University. MEEC is an annual, migratory conference organized by and for graduate students and postdocs. The conference is an excellent opportunity for students to network with others, learn about research being done in the Midwest, and present their own research in a relaxed and low-key environment. This year’s conference is particularly collaborative because of the proximity and involvement of the University of Missouri-St. Louis and Washington University. Registration by 15 January is $40; late registration until 30 January is $50. Information on registration, abstract submission, events, and field trips can be found at ‹http://wallace.slu.edu/meec2006/›. |

Instructions for ContributorsDEADLINES: Contributions for publication in the Bulletin must reach the Editor’s office by the deadlines shown below to be published in a particular issue: |
Issue
January (No. 1)
April (No. 2)
July (No. 3)
October (No. 4)Deadline
15 November
15 February
15 May
15 August
Please note that all material for publication in the Bulletin must be sent to the Bulletin Editor. Materials sent to any address except that of the Editor, given below, must then be forwarded to the Editor, resulting in delay in action on the manuscripts. Send all contributions, except those for Emerging Technologies, Ecology 101, Ecological education K–12, and Obituaries/Resolutions of Respect (see addresses below), to E .A. Johnson, Bulletin Editor-in-Chief, Department of Biological Sciences, University of Calgary, Calgary, Albert, T2N 1N4 Canada. Phone (403) 220-7635, Fax (403) 289-9311, E-mail: [email protected].MANUSCRIPT PREPARATION: The manuscript should be submitted as a WordPerfect or Microsoft Word (for Mac or DOS) manuscript, preferably as an e-mail message attachment to [email protected]. E-mailed photographs and diagrams must be in .tiff or .eps format. Other forms of electronic copy (text embedded in e-mail messages, diskettes sent by post) or hard copy can be submitted if absolutely necessary. If formatting could be troublesome (e.g., tables, European alphabet characters, etc.), hard copy also should be sent via fax to E. A. Johnson at (403) 289-9311, or via post. Hard-copy manuscripts should be double-spaced, with ample margins. Plain formatting must be used on hard-copy and electronic manuscripts. PLAIN FORMATTING consists of a single font of a single size, left justification throughout, line spacing the same throughout, and up to three different weights of headings. Other formats will not be accepted for publication. The author should THOROUGHLY PROOF the manuscript for accuracy, paying special attention to phone and fax numbers and web site and e-mail addresses, which are frequently incorrect.COVER PHOTOGRAPHS: The photo should illustrate ecological processes or an ecological research design. The cover of the July, 2004 issue is a good example. It helps if the colors in the photo are bright, although black and white photos are considered if they are well composed with good contrast.
If you would like to submit a digital file, submissions can be small jpegs (72 dpi) but if the image is selected for a cover the final image must be 300 dpi and at least 7 inches wide and 5 inches high. Email the file as an attachment to the Editor of the ESA Bulletin at [email protected]. Or send a single 5 x 7 or 8 x 10 photo to the Bulletin. On an accompanying photocopy, give your name, address, a photo legend up to 100 words, and, if the photo describes a paper in ESA or in another journal, the literature citation or title of the accepted manuscript. If you wish unused photos to be returned please include a self-addressed return envelope.LETTERS TO THE EDITOR AND COMMENTARIES: Please indicate if letters are intended for publication as this is not always obvious. The Bulletin publishes letters, longer commentaries, and philosophical and methodological items related to the science of Ecology. There are no page limits but authors may be asked to edit their submissions for clarity and precision. Previously published items from other sources can be republished in the Bulletin if the contributor obtains permission of the author and the copyright holder, and clearly identifies the original publication.MEETING ANNOUNCEMENTS: Submit a brief prose description of the upcoming meeting, including title, a short paragraph on objectives and content, dates, location, registration requirements, and meeting contact person’s name, street address, and phone/fax/e-mail address. Please do not submit meeting brochures in the expectation that the Editor will write the prose description; he won’t. Compare the publication deadlines above with the meeting deadlines to be sure the announcement will appear in time.MEETING REVIEWS: The Bulletin publishes reviews of symposia and workshops at the annual ESA meeting, as well as important and appropriate meetings that are unrelated to the annual ESA meeting. The reviewer should strive for a synthetic view of the meeting or symposium outcome, i.e., how the various presentations fit or conflict with each other and with current scientific thought on the topic. Review length is open, although about four double-spaced pages should be enough to capture the essence of most meetings.
The following advisory items are provided to help focus your review.
a) Meeting title, organizer, location, sponsoring organizations?
b) What were the meeting objectives, i.e., what scientific problems was the meeting organized to solve? Who cares (i.e., what was the relevance of this scientific problem to related ones under examination)?
c) How well did the meeting meet the objectives? Were there specific papers delivered or roundtables/discussion groups that were exemplary in reaching the objectives? You may concentrate the review on only the outstanding papers to the exclusion of all others, or give a comprehensive view of all presentations/meeting activities, or examine a selection of papers that neither describes all, nor focuses on a very few.
d) What new was discussed? What previously weak hypotheses were strengthened, confirmed or supported? Were any breakthroughs, or new or innovative hypotheses presented, that forced participants to rethink current concepts?
e) Was there anything else important that the meeting accomplished that may not have been part of its explicit objectives?
f) What subjects relevant to the meeting objectives were missing or left out? Did the scientific components of the problem that were included produce a strong slant or serious void by virtue of blind spots by the organizers, failure of invitees to appear, or similar difficulties?
g) Are there plans for a proceedings issue or meeting summary document, and if so who is editing it, who is publishing it, and when is it planned to appear (i.e., where can interested folks learn more about the meeting?)EMERGING TECHNOLOGIES: Submissions for this section should be sent to the Section Editors in charge of the section: Dr. David Inouye, Department of Biology, University of Maryland, College Park, MD 20742. E-mail: [email protected]; or Dr. Sam Scheiner, Div. of Environmental Biology, Natl. Science Foundation, 4201 Wilson Blvd., Arlington, VA 22230. E-mail: [email protected]ECOLOGY 101: Submissions should be sent to the Section Editor in charge of this section: Dr. Harold Ornes, College of Sciences, SB 310A, Southern Utah University, Cedar City, UT 84720. E-mail: [email protected]ECOLOGICAL EDUCATION K–12: Correspondence and discussions about submissions to this section should be sent to Susan Barker, Department of Secondary Education, 350 Education South,, University of Alberta, Edmonton, Alberta T6G 2G5 Canada. E-mail: [email protected]
(780) 492 5415 Fax: (780) 492 9402
or
Charles W. (Andy) Anderson, 319A Erickson Hall, Michigan State University, East Lansing, MI 48824 USA. E-mail: [email protected]
(517) 432-4648 Fax: (517) 432-5092FOCUS ON FIELD STATIONS: Correspondence and discussions about submissions to this section should be sent to E. A. Johnson, Bulletin Editor-in-Chief, Department of Biological Sciences, University of Calgary, Calgary, Albert, T2N 1N4 Canada. Phone (403) 220-7635, Fax (403) 289-9311, E-mail: [email protected].OBITUARIES AND RESOLUTIONS OF RESPECT: Details of ESA policy are published in the Bulletin, Volume 72(2):157–158, June 1991, and are abstracted below. The death of any deceased member will be acknowledged by the Bulletin in an Obituary upon submission of the information by a colleague to the Historical Records Committee. The Obituary should include a few sentences describing the person’s history (date and place of birth, professional address and title) and professional accomplishments. Longer Resolutions of Respect, up to three printed pages, will be solicited for all former ESA officers and winners of major awards, or for other ecologists on approval by the President. Solicited Resolutions of Respect will take precedence over unsolicited contributions, and either must be submitted to the Historical Records Committee (see ESA website) before publication in the Bulletin.









 (1)
(1)

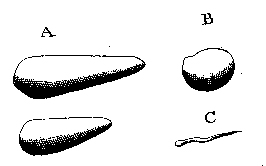
 Fig. 6. Now called Volvox, illustrating Leeuwenhoek’s letter of 2 January 1700 (Royal Society of London Philosophical Transactions 22:facing p. 483).
Fig. 6. Now called Volvox, illustrating Leeuwenhoek’s letter of 2 January 1700 (Royal Society of London Philosophical Transactions 22:facing p. 483). 
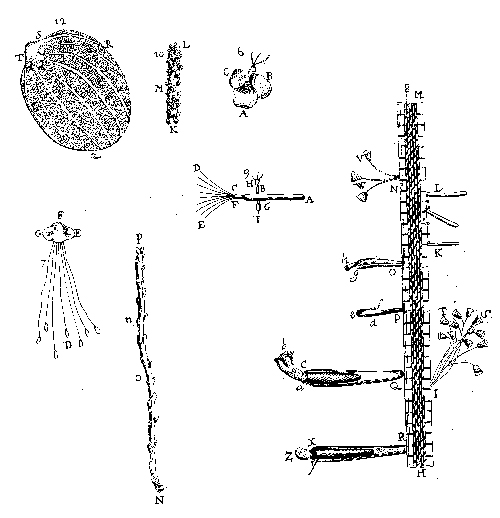
 a 6,000-hectare experimental forest operated by the State University of New York College of Environmental Science and Forestry (ESF) in the geographic center of the Adirondack Mountains (see the map on right).
a 6,000-hectare experimental forest operated by the State University of New York College of Environmental Science and Forestry (ESF) in the geographic center of the Adirondack Mountains (see the map on right). 